
Plant hormones
Hormones are organic molecules that can influence the physiology and development of plants and animals even at low concentrations. Hormones play an important role in the growth and flowering of the plant and many other things. This article briefly explains how plant hormones work in plants and how hormones ensure that plants flower.
Hormones are produced and transported throughout the entire plant. Simply put, they are signals, chemical signals, that can be sent and received throughout the entire plant. A leaf can and will transmit a signal to the end of the stem telling it to form flowers for example. The most well-known plant hormones are auxin, gibberellin, cytokinin, ethylene, and abscisin (abscisic acid). In addition, it has been demonstrated that brassino-steriods, salicylates, and jasmonates also function in a similar way to hormones. Hormones can also occur bonded to sugars or amino acids. In this form, they are inactive and provide storage. The hormones can be released again and become active under various conditions such as the influence of gravity or light for example.
Auxin
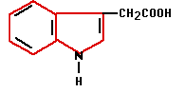 In the 1880s Charles Darwin and his son Francis started experiments that finally confirmed the existence of plant hormones. They experimented with oats and the influence of light on the direction of growth. Auxin was the plant hormone whose processes were demonstrated during these experiments. Auxin is produced in the plant’s growing points both above the ground and in the roots. Auxin influences water absorption, cell division, and cell stretching (it softens cell walls) among other things. Because auxin promotes the formation of roots on stems it is used in a variety of forms in rooting hormones.
In the 1880s Charles Darwin and his son Francis started experiments that finally confirmed the existence of plant hormones. They experimented with oats and the influence of light on the direction of growth. Auxin was the plant hormone whose processes were demonstrated during these experiments. Auxin is produced in the plant’s growing points both above the ground and in the roots. Auxin influences water absorption, cell division, and cell stretching (it softens cell walls) among other things. Because auxin promotes the formation of roots on stems it is used in a variety of forms in rooting hormones.
Experiments carried out by CANNA have shown that the effect of administering auxin depends very much on the concentration and method of application used for each plant type. With weak concentrations, flower formation is stimulated slightly and ripening takes longer. With high concentrations, there is an inhibiting effect on growth accompanied by deformities and tumour-like symptoms. Auxin that is produced in the tops of plants is capable of inhibiting the development of side shoots. This symptom is known as apical dominance. Removing the main tip stops the inhibiting effect and side shoots then develop which will eventually result in a broader plant. Where crop spacing allows only a few plants per square meter it is worthwhile removing the main tip as this makes it possible to use the light more efficiently. It’s also necessary to remove the tips regularly to achieve a good stock plant for propagation so that it will grow many more side shoots.
Gibberellin
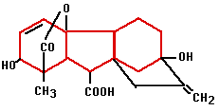 Gibberellin was first isolated in 1935 in Japan by Yabuta. The gibberellin was acquired from a fungus that had been the cause of reduced productivity for Japanese rice farmers for centuries. The gibberellin initially gave better growth but later in the season, it caused sterile fruits. Generally speaking, gibberellins work as growth accelerators because of cell stretching and cell division. They ensure that seeds germinate and flowers form in plants that need long days. Gibberellin is often used in the cultivation of fruit to help unfertilized pears and apples develop fully.
Gibberellin was first isolated in 1935 in Japan by Yabuta. The gibberellin was acquired from a fungus that had been the cause of reduced productivity for Japanese rice farmers for centuries. The gibberellin initially gave better growth but later in the season, it caused sterile fruits. Generally speaking, gibberellins work as growth accelerators because of cell stretching and cell division. They ensure that seeds germinate and flowers form in plants that need long days. Gibberellin is often used in the cultivation of fruit to help unfertilized pears and apples develop fully.
Administering gibberellin to short-day plants, or autumn flowerers, as they are also known, very quickly gives clear effects even at low concentrations. Plants become light green in color and stem split open because of the fast growth (photo 1). The plant’s speed of growth can reach 10 cm per day! Administering gibberellin during the vegetative phase causes plants to start flowering more slowly. Gibberellin is for short-day plants as testosterone is for people. It stimulates the formation of typically male organs and longer plants; longer internodes and male flowers in dioecious plants. When the pollen from these flowers is used to fertilize female flowers, seeds are created that always produce female plants.
Certain environmental influences can also cause the production of extra gibberellin. Plants will make more gibberellin in poorly lighted conditions, which causes them to become long and look lanky. Another effect is seen when the lamp is too close to the plant. Buds that are flowering can start to shoot again if the lamp is too close. This will cause the tops to become long and thin. To prevent this, the distance from the plant to the lamp during flower formation must be at least 50 cm for a 600W lamp.
Cytokinin
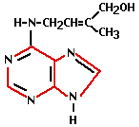 Cytokinin activity was first demonstrated in 1913. 30 years later it was discovered that a natural substance present in coconut milk was capable of helping plant cells multiply. Cytokinin was the responsible hormone for this. Cytokinin is known as the hormone responsible for cell division. It stimulates the metabolism and the formation of flowers on side shoots and as such is a counterpart to auxin. The cytokinin concentration is highest in young organs (e.g. seeds, fruits, young leaves, and root tips). High concentrations in an organ or tissue will stimulate the transport of sugars to those tissues or organs.
Cytokinin activity was first demonstrated in 1913. 30 years later it was discovered that a natural substance present in coconut milk was capable of helping plant cells multiply. Cytokinin was the responsible hormone for this. Cytokinin is known as the hormone responsible for cell division. It stimulates the metabolism and the formation of flowers on side shoots and as such is a counterpart to auxin. The cytokinin concentration is highest in young organs (e.g. seeds, fruits, young leaves, and root tips). High concentrations in an organ or tissue will stimulate the transport of sugars to those tissues or organs.
Administering cytokinin leads to greater leaf surface area and faster flower formation. However, the time that flowering finishes is comparable to untreated plants. Cytokinin can be seen as a counterpart to gibberellin in this regard because it stimulates the formation of female flowers on male plants.
Ethylene
 The practical use of ethylene comes from the time of Old Egypt when figs were scored to make them ripen faster. In 1934 it was discovered that plants produce ethylene themselves, which enables them to regulate fruit ripening. Ethylene is the least complex plant hormone from the molecular point of view and is produced by all organs. It is a gaseous hormone that is transported via the spaces between plant cells. It is responsible for fruit ripening, inhibition of growth, and leaf abscission (shedding). Ethylene has a stimulating effect on flower formation with certain types of plants (i.e. pineapples, mangoes, and lychees). Administering ethylene results in smaller plants and flowering finishes a lot quicker. The flowers ripen too quickly and consequently remain small.
The practical use of ethylene comes from the time of Old Egypt when figs were scored to make them ripen faster. In 1934 it was discovered that plants produce ethylene themselves, which enables them to regulate fruit ripening. Ethylene is the least complex plant hormone from the molecular point of view and is produced by all organs. It is a gaseous hormone that is transported via the spaces between plant cells. It is responsible for fruit ripening, inhibition of growth, and leaf abscission (shedding). Ethylene has a stimulating effect on flower formation with certain types of plants (i.e. pineapples, mangoes, and lychees). Administering ethylene results in smaller plants and flowering finishes a lot quicker. The flowers ripen too quickly and consequently remain small.
Because plants can be very sensitive to ethylene, the concentration is expressed in parts per billion parts of air (ppb). Concentrations of just 10 ppb can cause abnormalities in tomatoes. In situations where ripening flowers come in contact with young plants, there is the risk of accelerated ripening in the young plants. The ethylene that is produced can reach the young plants via the air. Ventilating occasionally (once per day) will remove the ethylene that has formed. High concentrations cause leaves to turn yellow immediately.
Ethylene can also accumulate around roots if they are wet for too long. This can lead to leaf chlorosis, stem thickening, leaves bending towards the stem, and greater susceptibility to diseases.
In stress situations, for example, when there is disease present or damage to the plant, the plant produces more ethylene, which causes it to remain smaller and finish flowering faster. Mechanical stress such as air movement can also cause the plants to produce extra ethylene, which will result in smaller plants with thick, sturdier stems. When the fans are too close to the plants there will be too much stress and this will adversely affect the yield.
Abscisin
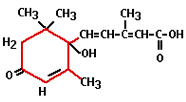 Abscisin was first isolated in 1963 and has the Latin word abscissio (breaking off) to thank for its name. This is because people thought that abscisin was responsible for the breaking off (shedding) of leaves and fruits, however, it was later shown that ethylene plays a much more direct role in this.
Abscisin was first isolated in 1963 and has the Latin word abscissio (breaking off) to thank for its name. This is because people thought that abscisin was responsible for the breaking off (shedding) of leaves and fruits, however, it was later shown that ethylene plays a much more direct role in this.
Abscisin is produced in the chloroplasts of older leaves and has both inhibiting (growth) and stimulating (protein storage) characteristics. When there is a large supply of abscisin to the growing points of the stem and roots, cell division stops and the plant enters a rest period.
Abscisin is an important hormone as far as stress situations are concerned. It is responsible for closing the stomata when the plant is under water stress due to continuing high temperatures, low atmospheric humidity, or an EC in the feeding medium that is too high.
Flower formation in short-day plants
Even though a lot of research has already been done into the changeover from growth to flowering in plants, it still hasn’t been explained how this mechanism works exactly. In the case of short-day plants, the formation and development of flowers depend on the precise length of the night. Short-day plants will flower when the nighttime period is longer than 12 hours. It is important that it is really dark during this period because the plant is only capable of measuring the period of darkness and not the period of light. Almost any light level during the dark phase will affect the cycle. This is measured in the leaves, which then send a signal to the furthermost ends of the branches instructing them to form flowers. The hormone that gives this signal is called florigen. So it is theoretically possible, for example, to use material from flowering plants to stimulate other plants to flower under 18 hours of light.
Different hormones play an important role in the phase following the first set of flower buds. So cytokinin and auxin play an important role in the further formation and growth of the flowers. Abscisin and ethylene are important during ripening.
Using hormone preparations
If you want to experiment with plant hormone preparations, pay close attention to how, when, and how much hormone you use. The final effect depends on many factors such as the time of administering (which phase, time of the day), the route chosen for administering (leaf or roots), and the concentration. The final effect of administering hormones can depend very much on the concentration used. For instance: weak concentrations of Auxin stimulate root growth while strong concentrations cause extra ethylene production, which, in turn, causes the plant to finish flowering faster.